The ability of a cow or heifer to successfully mate, conceive, give birth, and raise a healthy calf each year is essential for profitable and sustainable beef production. A good understanding of anatomy and physiology of both the male and female is helpful in successfully managing reproduction.
Causes for failures in reproduction must be identified and overcome. Research has led to the development of numerous techniques for managing the reproductive processes of animals. These techniques provide many options to help progressive cattle producers accomplish their production goals.
Anatomy
Ultimately, reproductive processes are regulated at the level of the brain. The hypothalamus controls several body processes and behaviors along with reproductive processes. Body temperature, concentration and components of body fluids, and the drive to eat and drink are just a few functions of the hypothalamus. It is classified as a neuroendocrine gland since it sends and receives neural signals through the nervous system and hormonal messages through the endocrine system.
The second organ, the pituitary gland, sits at the base of the brain. The pituitary is about half an inch in diameter and weighs about 1 gram. Physiologically, the pituitary is divided into two distinct regions: the anterior and posterior pituitaries. Each region secretes various hormones that direct body processes. Some of these hormones are responsible for reproductive events, while others control growth, metabolism, and water balance.
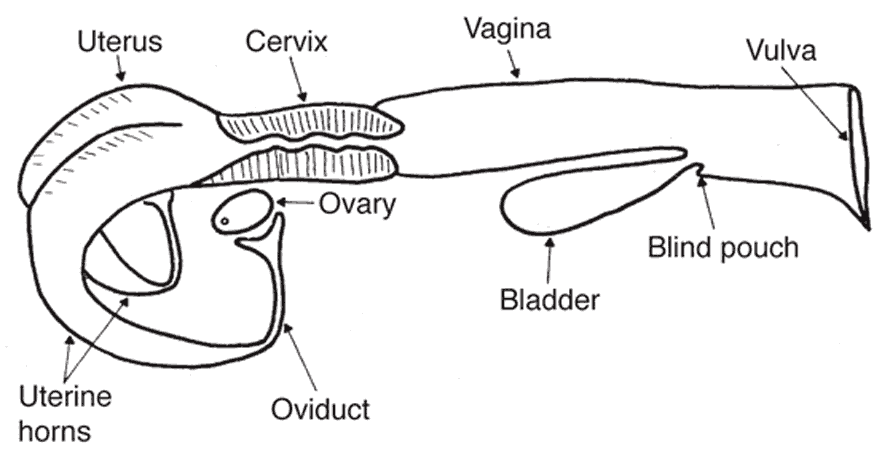
The female reproductive organs consist of the ovary, uterus, cervix, vagina, and vulva. A sketch of the reproductive tract of the cow is shown in Figure 1. Female reproductive tracts of various livestock species are similar to the tract of the cow but differ primarily in the shape of the uterus and cervix.
The oviduct, also known as the Fallopian tube, begins as a funnel-shaped tube that engulfs the ovary. This funnel portion of the oviduct is called the infundibulum. When ovulation occurs, the oocyte is picked up by the infundibulum and channeled into the oviduct. Fertilization takes place in the oviduct if viable sperm are present, but the oocyte remains capable of fertilization for only a short time. Therefore, it is essential that sperm be present in the oviduct near the time of ovulation. The oocyte moves through the oviduct into the uterine horn within the next 3 to 4 days. If the ovum is fertilized, it begins embryological development; if not, it simply degenerates.
The body of the uterus of the cow is short, while the uterine horns are relatively long and well developed. The fertilized embryo moves from the oviduct into the uterine horn, where fetal and maternal membrane development begins. This newly developing fetus grows within a layer of membranes called the placenta, through which nourishment from the dam diffuses. This point of interface is called the placentome. It is made of the joining of the caruncle (maternal) and cotyledon (fetal). There is no direct blood connection between the fetus and the dam in the bovine placenta, but rather a complex system that selectively allows certain molecules to pass from the maternal side of the placenta to the fetal side and vice versa. It also provides nutrients and carries waste products from the fetus.
The cervix is a denser structure that contains more connective tissue. The bovine cervix is often described as having a “turkey neck” feel when palpated. It has thick walls and a small opening, or cervical os. The cervix softens and relaxes around the time of estrus, the period of sexual receptivity. Cervical softening allows a passageway for sperm at mating and also for expulsion of the fetus at the time of birth. During pregnancy, the cervix is filled with a thick mucous secretion known as the cervical plug. This provides a physical barrier between the vagina and the uterus, in order to protect placental structures and the developing fetus. In the days prior to calving, the cervical plug is expelled, and the cervical opening begins to dilate.
The vagina serves as a receptacle for the male’s penis during natural mating. In natural mating, semen is deposited in the vagina near the cervix. In artificial insemination, a catheter is inserted in the vagina and manipulated through the cervix, allowing a smaller number of sperm cells to be deposited on the uterine side of the cervix. Urine is discharged from the urinary bladder through the urethra, which opens into the base of the vagina. The region behind the urethral opening is called the vestibule and is a common passageway for both the urinary and reproductive systems. The external opening of the vagina is called the vulva.
Physiology
The ovary produces the oocyte by a process called oogenesis. In contrast to spermatogenesis in the bull, which is continuous, oogenesis is cyclic. This cycle of oocyte development in cattle is called the estrous cycle and is typically 21 days in length. During the estrous cycle, two prominent structures are present within the ovary: the follicle and corpus luteum.
Each of these structures undergo a development and subsequent regression phase during the estrous cycle. Follicles begin within the ovary as one of several thousand primary follicles, which consist of a germ cell surrounded by a layer of flattened cells. This germ cell has the potential to mature into an oocyte if the follicle completes the development phase. However, only a small percentage of primary follicles continue through the secondary and tertiary follicular phases and ultimately undergo ovulation. Primary follicles that never complete development undergo atresia and are replaced by newly formed primary follicles.
The relatively few primary follicles that complete development do so through a series of phases. Many layers of cells are added to the single layer of cells surrounding the oocyte in the primary follicle and a central cavity forms. The follicle and cavity within the follicle grow larger, and the oocyte becomes attached to a stalk of cells on the side of the follicle opposite the future site of ovulation. As the follicle continues to grow larger, the outer layer of the follicle becomes thinner. This follicle is mature and called a dominant follicle. The outer layer of the follicle ruptures at the appropriate time, and the oocyte and contents of the follicular cavity are released. Follicular development occurs in concert with other reproductive and behavioral functions so that, near the time of ovulation, the female reproductive tract is prepared to receive both the oocyte from the female and sperm from the male.
Following ovulation, the cells that developed within the follicle undergo a differentiation process. This process is called luteinization and gives rise to the second prominent ovarian structure, the corpus luteum (pl. corpora lutea). This structure is often referred to simply as the CL and has the important function of secreting the hormone progesterone.
The CL goes through a maturation and regression cycle much the same as the follicle. A blood clot-type structure known as a corpus hemorrhagicum forms in the cavity left by the ruptured follicle and is transformed into a CL by Day 5 of the cycle (Day 0 = estrus). The CL is fully functional from Day 5 to Day 15 of the cycle and then begins to regress if the female does not become pregnant. The CL regresses and no longer secretes progesterone as the follicle of the next estrous cycle begins to develop. As the CL regresses further, it becomes known as the corpus albicans and remains visible on the ovary for several subsequent cycles.
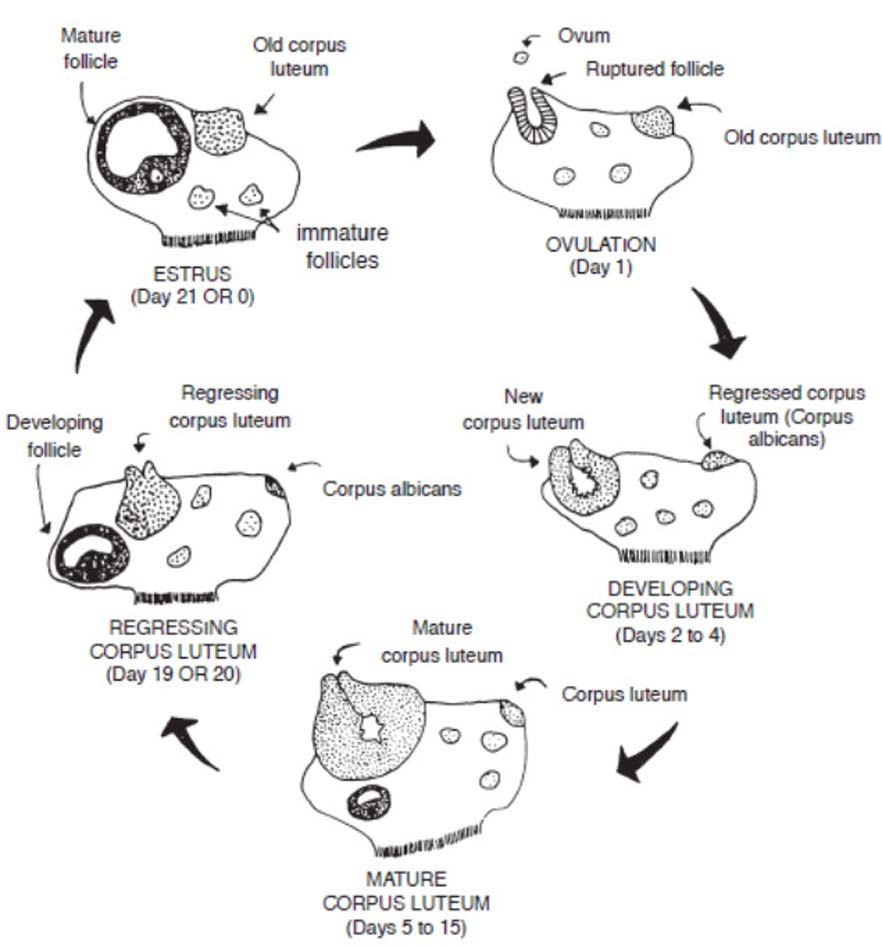
Figure 2 illustrates the changing structures on the ovary during a typical 21-day estrous cycle. The dynamic development and regression of the corpus luteum and follicles are a continual process in the normal cycling cow until she becomes pregnant. In a pregnant female, CL regression does not occur, and the cyclic activity stops until after calving. A cow generally remains anestrous (does not cycle) for a period of time following calving. The length of the postpartum anestrous period is on average around 60 days, but this is highly variable among cows based on age, nutrition, lactation, presence of a suckling calf, environmental stress, and numerous other factors. Management to control the length of this interval is a main consideration in successful cow-calf enterprises, since a cow must rebreed within 85 days after calving to produce a calf on a yearly interval.
Any condition that prolongs the period of time that blood levels of progesterone remain high will have the same effect as pregnancy in stopping the regular 21-day interval between occurrences of estrus. Occasionally the CL does not regress normally (persistent CL) even though the animal does not become pregnant. This requires diagnosis and treatment by a veterinarian.
Abnormally short estrous cycles (7 to 11 days) can occur. This condition appears to be caused by either no corpora lutea being formed, or if one is formed, it is non-functional and progesterone levels remain low. An estrous cycle can be shortened intentionally by administering a hormone called prostaglandin F2a, which causes the CL to regress. Prostaglandin F2a is a key tool used in estrus synchronization protocols
Estrus is not always accompanied by ovulation, nor is ovulation always accompanied by estrus. Heat without ovulation (anovulatory heat) will not result in pregnancy even though the female may be serviced. Ovulation without behavioral estrus (silent heat) is not uncommon in cows, especially the first few weeks after calving. Such females will not accept service from a male, but artificial insemination can result in pregnancy if semen is placed at the appropriate time in advance of ovulation.
There are wide differences between mammalian species in various aspects of the estrous cycle. The cow is in a group of species that exhibits estrus more than one time per year and is therefore called polyestrus. Even though cows will cycle at any time during the year, research suggests day length may influence cow fertility, especially in some breeds or biological types. Slightly lower fertility has resulted during the short-day length period of the year as compared to the long-day length period of the year.
The hormones of female reproduction
Reproduction in the female is controlled by numerous hormones secreted from specialized glands called endocrine glands. These secretions are produced in the glandular cells and pass into the blood and lymph systems for transport to specific parts of the body where they produce their function.
The hormone, estradiol (or estrogen) is produced by the granulosa cells of the follicle. Estradiol has various effects, including:
- Development and function of the secondary sex organs
- Receptivity to mating during and other behaviors associated with estrus
- Rate and type of growth, especially relative to deposition of fat
- Regulation of puberty attainment and length of the postpartum anestrous period
Progesterone, secreted by the corpus luteum, suppresses ovulation of a dominant follicle and the associated secretion of high levels of estradiol. Therefore, elevated levels of progesterone prevent a cow from coming into heat. Progesterone is necessary for preparing the uterus to receive the fertilized ovum and maintains the proper uterine environment for continuation of pregnancy.
Estradiol and progesterone are not separate in their functions since both must be present for certain processes to occur. For example, the ratio of the concentration of estradiol to progesterone dictates the onset and duration of behavioral estrus. Development of the uterus is initiated by estradiol and completed by progesterone. Estradiol causes contraction of the uterus near the time of estrus and ovulation, which aids in sperm transport. Progesterone has a quieting effect on the uterus so that there are no contractions which might disturb pregnancy.
The production of ovarian hormones is under direct influence of gonadotropic hormones produced by the anterior pituitary. Follicle stimulating hormone (FSH) and luteinizing hormone (LH) are secreted from the pituitary and travel through the blood to the ovary. The release of FSH and LH is mediated by gonadotropin-releasing hormone (GnRH) by the hypothalamus. FSH stimulates the recruitment, growth, and development of follicular waves, while LH supports continued development of a dominant follicle. Additionally, a surge in LH release around the time of onset of estrus causes the follicle to initiate the ovulatory process and develop into a corpus luteum.
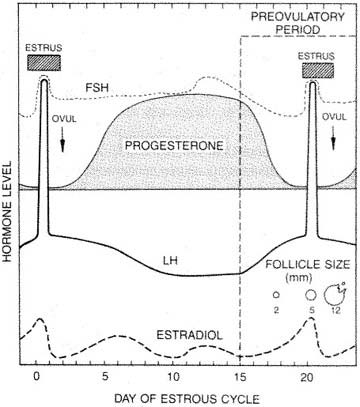
The cyclic rise and decline of the reproductive hormone concentrations is graphically illustrated in Figure 3. This cyclic process continues every 21 days in a normal-cycling cow, but changes if conception occurs. Following pregnancy and a period of anestrus, the estrous cycle once again continues. Figure 3 shows the female hormones during a typical estrous cycle.
The estrous cycle
The reproductive cycle of the cow consists of a series of events that occur in a definite order over a period of days. In the cow, this cycle averages 21 days in length, although this can range from 17 to 24 days. In this cycle, the reproductive system is prepared for potential establishment and maintenance of pregnancy. Alternatively, in the event that fertilization does not occur or that establishment or maintenance of pregnancy fails, the female returns to estrus. Figures 2 and 3 show the ovarian changes and sequence of events in a typical 21-day cycle in which pregnancy does not occur.
- Days 0 to 1
The cow is in for estrus (standing heat) on Day 0 for an average of 18 hours (range 8 to 24 hours). Shortly after onset of estrus, a surge of LH is released by the pituitary gland. About 30 hours after the start of the standing heat, the mature dominate follicle ruptures (ovulates). - Days 1 to 2
The cells that formerly lined the follicle change and become the luteal cells of the corpus luteum. This change in cell forms is caused by hormonal action, primarily that of LH. - Days 2 to 5
The corpus luteum grows rapidly in both size and function. Numerous follicles may be seen on the ovary at this stage, but by Day 5 they have begun to regress. - Days 5 to 16
The corpus luteum continues to develop and reaches its maximum growth and function about Day 10. It secrets the hormone progesterone, which inhibits (blocks) LH release by the pituitary gland. During this period, the ovaries are relatively inactive except for the functional corpus luteum and wave-like follicular development. A dominant follicle will reach maturity but the existence of the high levels of progesterone inhibit estrus and ovulation. - Days 15-17
The corpus luteum regresses rapidly due to luteolytic activity of the uterus caused by the release of the hormone, prostaglandin F2a. - Days 18 to 2
The corpus luteum is almost non-functional and this releases the blocking action of progesterone. Of the several follicles that commence growth, one becomes more prominent by a surge in rapid growth and activity. As the dominate follicle grows, it secretes increasing amounts of estradiol. The remainder of the follicles regress. - Day 21 or 0
With the increase in estradiol release by the dominant follicle and a corresponding decrease in progesterone by the regressing corpus luteum, estrus or heat will occur (cycle has now returned to Day 0). The high estradiol level in the blood triggers a release of LH near the end of heat. Following this surge in blood levels of LH, the mature follicle ruptures to release the ovum and the cellular tissue left behind becomes luteinized in response to the stimulation of a hormonal complex to form a new corpus luteum (cycle has now returned to Days 1 and 2). Progesterone again becomes the dominant hormone.
It should be noted that the timing given for the preceding events is only approximate and differs from female to female based on cycle length and the number of follicular waves per cycle. It should also be noted that, if fertilization occurs, and the maternal recognition process begins by the release of interferon tau, the corpus luteum does not regress but continues to function by secreting progesterone for maintenance of pregnancy.
Portions of this guide were adapted from Nebguide G80-537, by G.H. Deutscher, Extension Beef Specialist, University of Nebraska.