Understanding how plants function (physiology) and their form and structure (morphology) is important when managing grazing animals because both physiological and morphological changes affect plant survival and the quantity and quality of forage. An awareness of how plants respond physiologically and morphologically to their environment provides the basis for understanding how plants react to external factors such as high and low temperatures, drought, light and grazing.
This publication presents an overview of basic plant physiology and morphology, and discusses the impact of form and function of plants on grazing management. For more information on the topics discussed in this publication, see the sources listed under Additional reading.
Photosynthesis
Green plants are able to trap and store energy from sunlight. Photosynthesis occurs in the chloroplasts of green plant tissue. Plants make simple sugars, or carbohydrates (CH2O), and release oxygen (O2) using light energy from the sun, carbon dioxide (CO2) from the air, and water (H2O) from the soil (Figure 1). This reaction supports all life on earth. The carbon of CO2 is converted to the carbon of carbohydrate and is a chemical way to store the sun’s energy as food. Because CO2 is not abundant (0.036 percent in the atmosphere), the success of a plant depends on its ability to collect and use CO2.
Plant tissue consists of complex carbohydrates synthesized from simple carbohydrates. However, energy production in a sward (herbage in a pasture) is a relatively inefficient process. Only 2 to 6 percent of the sun’s energy that reaches a sward is converted into plant growth. Energy accumulation in a sward is determined by the amount of light intercepted by the leaf canopy and the efficiency of the photosynthetic process. Light interception depends on the amount of leaf in a sward, which is expressed as leaf area index (LAI). LAI is the surface area of leaf blades per unit area of ground. Virtually all of the light is intercepted when LAI reaches values of about 6 for grass pasture, 4 for grass-legume mixtures and 3 for white clover pastures.
The structure and morphology of a sward can have other effects on photosynthetic efficiency. For example, young leaves have higher efficiency than older leaves, and grass leaves developed in high-intensity light have a higher efficiency than those grown in the shade. Therefore, the need to maintain a sward with a high LAI must be balanced against the need of young leaves to be exposed to direct sunlight.
Cool-season (C3) and warm-season (C4) plants have different photosynthetic systems. Cool-season plants originated from temperate regions and have a C3 system, whereas warm-season plants evolved under tropical conditions with the C4 photosynthetic pathway. Warm-season plants are more efficient than cool-season plants when both are grown under optimal conditions because the C4 system is more efficient than the C3 system at gathering CO2.
The optimal temperature for growth is 65 to 75 degrees F for cool-season plants and 85 to 90 degrees F for warm-season plants. Warm-season plants use a little more than half as much water as cool-season plants to produce the same amount of dry matter. Because warm-season plants have higher water-use efficiency and higher optimum temperature for growth, warm-season pastures are more productive during the hot, dry summer months. Conversely, cool-season pastures are more productive in the cooler, more moist spring and fall months.
The points outlined above explain why cool-season grasses grow in the spring and mature by late spring or early summer, then become semidormant in the summer and resume growth in the autumn, and why warm-season grasses begin growth in late spring, mature during late summer and become dormant by early fall.
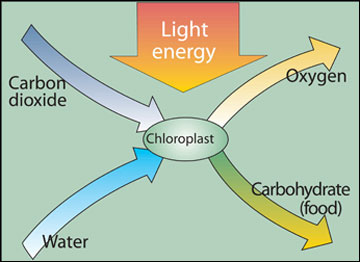 Figure 1. Photosynthesis takes place in the chloroplast of the cells of green plant tissue. Carbon dioxide enters the leaves through stomata in the leaf surface and passes into the cells by diffusion. Water is transported to the leaves from the roots.
Figure 1. Photosynthesis takes place in the chloroplast of the cells of green plant tissue. Carbon dioxide enters the leaves through stomata in the leaf surface and passes into the cells by diffusion. Water is transported to the leaves from the roots.
Carbohydrates
Through photosynthesis, the green tissue of plants is able to manufacture carbohydrates. Plants only produce carbohydrates during the day and during the growing season. When photosynthesis exceeds immediate carbohydrate (CHO) needs, excess carbohydrates are stored in the roots of legumes or stem bases of grasses. Carbohydrates are used to maintain the plant and for growth after defoliation or dormancy. Plants use stored carbohydrates to survive during winter and, when conditions become more suitable, to renew growth. Carbohydrates can be stored in storage organs such as stem bases, roots, rhizomes and stolons. Storage of excess carbohydrates is vital to the survival of the plant because carbohydrates are the only source of energy for new growth after dormancy, severe defoliation or environmental stress.
Growth has priority over storage for carbohydrate use. Carbohydrate storage increases when growth rate declines and leaf area is large. In contrast, carbohydrate storage decreases when the growth rate is fast and leaf area is small. When the plant produces more carbohydrates than are needed for growth and maintenance, the excess is moved to the storage organs. Frequent and close defoliation can totally deplete stored carbohydrates. Plants must be allowed the opportunity to store carbohydrates for those times when photosynthesis is unable to produce enough for growth.
The grass tiller
A single grass shoot is called a tiller (Figure 2). An individual plant is made up of many tillers, and a typical pasture has about 30 plants and 400 to 1,000 tillers per square foot. A tiller typically has three to four visible leaves, and the lowest leaf is the oldest. Moving up a tiller from the bottom, each leaf is younger than the one below, and successive leaves are on opposite sides of the tiller. Each leaf has an upper part called the blade and a lower part called the sheath. Leaf sheaths could be rolled into a structure called a pseudostem. Below the pseudostem is a true stem. The true stem has divisions called nodes. A shoot bud is found at each node, and roots also emerge near the nodes. The length of stem between the nodes is called the internode. The “growing points” of grass are found just above the node of each stem.
The tiller is essentially a single growing point encased in the sheaths of leaves that grow from it. As it grows, the tiller develops its own root system and the capacity to develop new generations of tillers from buds at the base of individual leaves. Although a grass plant will have many generations of individual tillers and each tiller is self-supporting, the original connection between tillers is usually not severed. This connection allows the transfer of nutrients between tillers, particularly following defoliation.
An individual tiller (shoot) is composed of a growing point (apical meristem), a stem, leaves, roots, nodes (joints) and dormant buds. Depending on the species, buds may be located at the base of the tiller or nodes, on the stem, and/or at the nodes on stolons or rhizomes (Figure 2). Each of these dormant, or inactive, buds has the potential to produce a new tiller.
All tillers begin as a growing point developing from a dormant bud at or below ground level. Vegetative tillers can produce an infinite number of leaves. However, once a tiller receives the message to become reproductive and the tiller stem elongates between the nodes (on the internode), no more leaves can be initiated. Between vegetative and reproductive tiller development, the tiller appears elongated although no seed head is apparent. In some grasses, tillers may remain elongated (jointed) as vegetative tillers. The transition phase is followed by the reproductive stage, which is evidenced by the emergence of the seed head (inflorescence). Removing the reproductive tiller breaks the dormancy of the buds associated with that tiller. Following the removal of a reproductive tiller, the most rapid growth will be from a tiller bud at the base of the defoliated tiller. The aerial tillers associated with some grasses, such as switchgrass and reed canarygrass, are the least productive of the new tillers.
Early in the growing season, all grasses have their growing points at or near ground level, which is below the height at which cattle can graze. About half the growing points of grasses develop into seed heads and are pushed above the canopy when the stem elongates. The remaining growing points of ryegrass, tall fescue, Kentucky bluegrass and many other grasses remain at or below ground level throughout early spring. In contrast, the remaining growing points of other grasses, such as smooth bromegrass, timothy, reed canarygrass and switchgrass, elongate the internodes and elevate the growing point relatively early in their development.
At certain times of the year, some grasses, such as smooth bromegrass and reed canarygrass, may have elongated growth during summer without seed head development. When a grass plant that has its growing point at or near ground level is grazed, new leaf material can be produced from the stem apex of the grazed tiller (Figure 3). However, when the growing point has been elevated and then is removed by grazing, new leaf material must come from tiller buds that have been dormant.
When new growth comes from a tiller bud, the energy required to get it started must be provided by either a residual leaf or stored carbohydrates. Unless some leaf material remains, the only source of energy for the developing bud is that stored in the plant’s storage organs. On a plant that is less severely defoliated, the developing bud can receive energy manufactured by the remaining leaf material, in which case the plant’s carbohydrate reserves are not depleted and do not have to be replenished. However, the plant will be harmed if it repeatedly has to use its stored carbohydrates because of close defoliation, as it will not have time to replenish carbohydrates between defoliations.
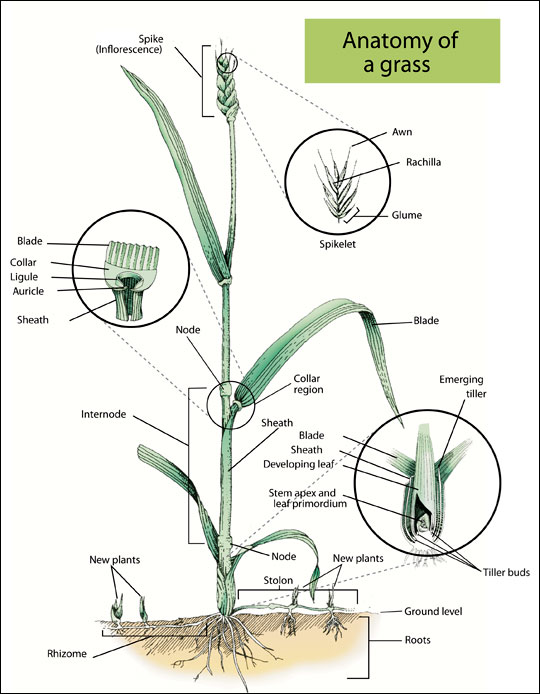
Figure 2. Although all grasses look somewhat different, this stylized drawing shows the major parts of a grass plant.
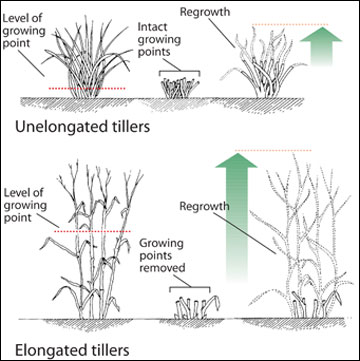 Figure 3. Regrowth varies among grasses. Grazed grass plants with growing points close to ground level (top) regrow differently than grazed grass plants with elevated growing points (bottom).
Figure 3. Regrowth varies among grasses. Grazed grass plants with growing points close to ground level (top) regrow differently than grazed grass plants with elevated growing points (bottom).
Root development
Like leaves, roots grow and die, but roots live much longer than leaves. Whereas a tiller may have three to four nodes with emerged leaves, it will have about 10 nodes with roots attached.
Of the total photosynthesis, about 15 percent is allocated to the root system. However, plants vary the proportion of photosynthate to roots and shoots during the season. Younger roots receive more photosynthate than older roots. During seed head development, allocation of photosynthate to roots is decreased. During late winter or after a summer drought, allocation to roots is temporarily increased. Following removal of the shoot, the levels of carbohydrates in the roots are low, and root growth can be suspended.
For some pasture plants such as tall fescue, leaving more herbage at grazing may allow the plant to better withstand a period of low water availability. Because removing only part of the canopy does not interfere with the continued development of the root system, the plant will have deeper roots. In a drought, a deep root system has access to a larger soil volume for water.
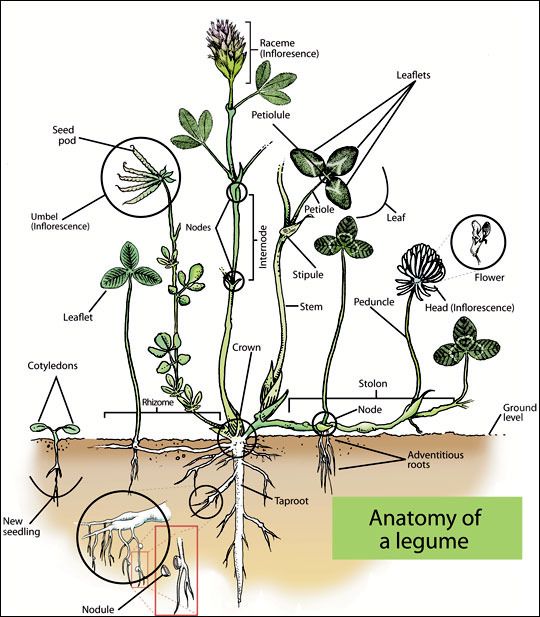
Figure 4. Although all legumes look somewhat different, this stylized drawing shows the major parts of a legume plant.
Summary
The leaves of plants take in carbon dioxide from the air through tiny pores. Using solar energy, the leaves recombine the carbon with oxygen and hydrogen to make sugars and carbohydrates. Minerals and nitrogen from the soil are combined with the sugars to make fiber, protein, plant oils and fats. The plants use the sugars, starches, proteins, oils and fats to grow and reproduce. Minerals from the soil make up about 5 percent of the dry weight of leaves, stems and roots. Carbon, hydrogen and oxygen from the air, and water from the soil make up most of the remainder.
The ability of some grasses and legumes to recover quickly after defoliation makes them valuable for forage production. However, removing too many leaves reduces forage production and damages the root system of the plant. Plants subjected to repeated, severe defoliations will soon die. Root growth is closely related to the production of forage. Plants maintain maximum root vigor and growth if no more than half their leaf area is removed at each grazing. Removing more than half of the plant will slow or even stop root growth. During the growing season, perennial plants store food (carbohydrates) in the bases of the shoots of grasses or the roots of legumes. The plants use these reserves to live while dormant and to make the first new growth after dormancy or defoliation.
The growing points of grass are located just above the uppermost node of each stem. Early in the season, the stem is very short and the growing points are located at the base of the plant. As the season progresses, the internodes of most species elongate and push upward to expose a seed head. These elongated tillers will produce no new leaves and, because of the stems, are of lower quality than vegetative tillers. Some species elongate relatively early in the growing season, and removal of these tillers means that new growth must come from the base of the plant. Plants of this type require a longer interval between defoliations so that the their root reserves can be replenished.
Additional reading
- Ball, D.M., C.S. Hoveland, and G.D. Lacefield. 1991. Southern forages. Potash and Phosphate Institute: Foundation for Agronomic Research: Altanta, Ga.
- Cherney, J.H., and D.J.R. Cherney, eds. 1998. Grass for dairy cattle. CABI Publishing: New York, N.Y.
- Hodgson, J. 1990. Grazing management: Science into practice. Longman Group UK Limited: United Kingdom.
- Hodgson, J., and A.W. Illius, eds. 1996. The ecology and management of grazing systems. CAB International: Wallingford, United Kingdom.
- Waller, S.S., L.E. Moser, and P.E. Reece. 1985. Understanding grass growth: The key to profitable livestock production. G.A. Gates. ed. Trabon Printing Co.: Kansas City, Mo.
This publication replaces Chapter 3, Growth of Forage Plants, in MU Extension publication M168, Dairy Grazing Manual. Original author: Greg J. Bishop-Hurley, University of Missouri.